A tale of two leaflets
A remarkable feature of the plasma membrane is that its two halves are chemically distinct: the lipids that face the internal contents of the cell are very different from the lipids that face the external environment. The plasma membrane that surrounds a cell has a distinctive compositional asymmetry, with sphingomyelin and PC lipids enriched in the outer leaflet and the aminophospholipids PS and PE enriched in the inner leaflet. In the field of membrane biophysics, a long standing goal is the preparation of liposomes that mimic this asymmetry.
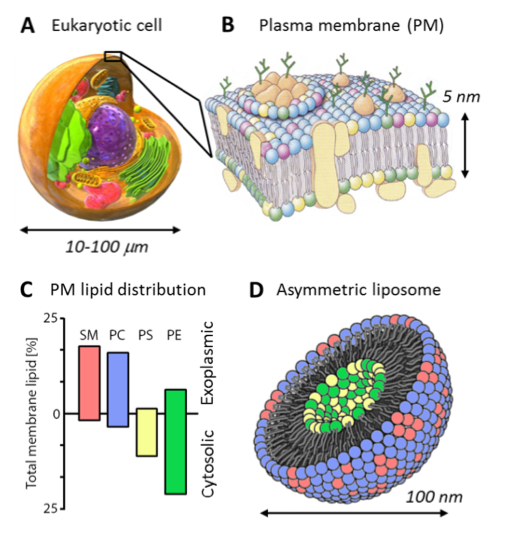
That cells actively segregate different types of lipids between the two leaflets has been known since the early 1970s. Nearly 50 years later, it is still not clear what role this compositional asymmetry plays in the life of a healthy cell, yet it is certain that cells do not function properly without it, and expend valuable energy to maintain it. For example, one important class of lipids called phosphatidylserines (PS) are located exclusively in the inner leaflet of healthy cells. This is due to the action of proteins called “flippases” that move them in one direction (from the outside to the inside). A buildup of PS in the outer leaflet is a signal that something has gone terribly wrong, and triggers special immune cells called phagocytes to eat the dying cell.
Mounting evidence suggests that the lipid compositional asymmetry of the plasma membrane not only confines certain lipids to their respective leaflets in order to facilitate their direct interaction with other molecules, but also results in unique membrane properties that, although currently poorly understood, are likely to be crucial for normal cellular function. Our research aims to shed light on the mystery of cell membrane asymmetry by answering the basic question of how the two halves of a membrane communicate with and influence each other. We achieve this through systematic study of the different types of lipids that are found on each side of an asymmetric membrane: how they influence the properties of the other half, as well as the properties of the membrane as a whole. And because much of a membrane’s functionality is derived from the proteins it hosts, we also investigate how membrane-resident proteins affect—and are in turn affected by—lipid asymmetry.
Improving the preparation of asymmetric liposomes
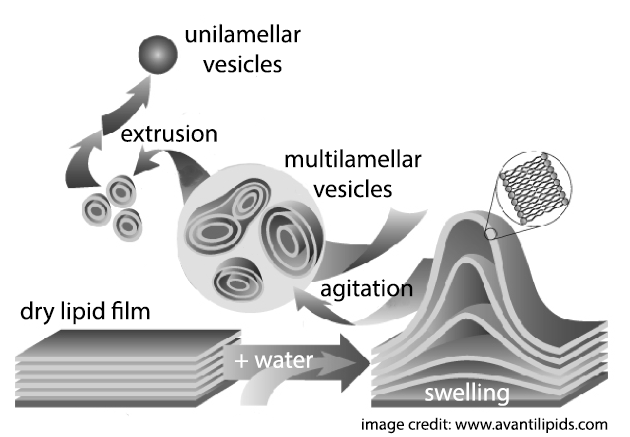
Lipid bilayer vesicles are among the most widely used model systems for studying membrane properties. Liposomes are typically prepared by drying a lipid film on the bottom of a glass vial and then hydrating it with buffer to produce multilamellar vesicles or MLVs, similar to onions in appearance (though 1000 times smaller!). The MLV suspension is then forced through a filter with microscopic pores of uniform diameter (typically 100 nm) in a process called extrusion. In addition to resizing the vesicles, the shearing forces generated by extrusion strip the layers from the onion-like MLV structures. After 20 or 30 passes through the extruder, what’s left is a fairly uniform population of ~100 nm diameter, single bilayer vesicles. Liposomes prepared in this time-tested manner are extremely versatile and can be made with nearly any lipid or mixture of lipids, allowing researchers to study a composition that mimics their favorite biological membrane. However, what this method cannot do is put different lipids on different sides of the bilayer! In other words, bilayers prepared by conventional methods are compositionally symmetric.
The ubiquity of membrane asymmetry in living cells hints at its importance for normal cell function. Symmetric model membranes prepared by conventional dry film hydration and extrusion almost certainly lack at least some key structural characteristics of natural cell membranes that may critically affect how those membranes interact with proteins and small molecules. There has long been a widely recognized need to expand the biophysical toolkit with model systems that more closely mimic the asymmetric cell membrane environment, while still allowing for easily controlled variation of the inner and outer leaflet lipid compositions.
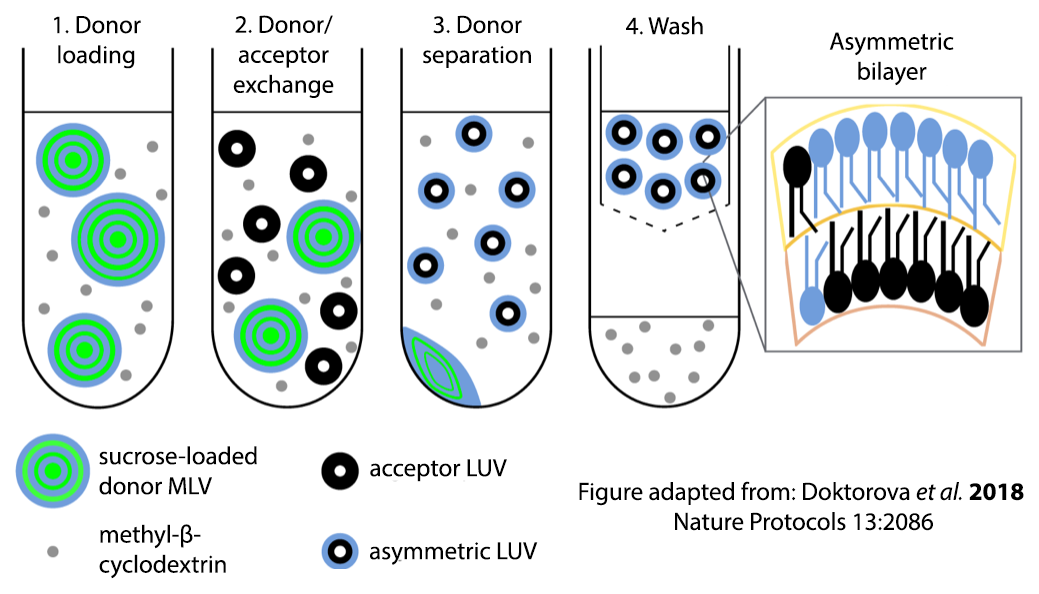
It is not easy to make an asymmetric membrane in a laboratory, and an important part of our research is the development and continual improvement of techniques for creating artificial membranes that mimic the lipid asymmetry found in real cell membranes. These liposomes are chemically far simpler than a living cell and therefore easier to study and understand at the level of individual molecules. Artificial membranes also have many other uses, including the encapsulation of therapeutic drugs for delivery to specific sites in the body. Better protocols for preparing highly reproducible asymmetric liposomes of precisely controlled composition may ultimately prove beneficial for these applications as well.
Research articles
Doktorova M, Heberle FA, Eicher B, Standaert RF, Katsaras J, London E, Pabst G, Marquardt D. 2018. Preparation of asymmetric phospholipid vesicles: The next generation of cell membrane models. Nature Protocols 13:2086-2101.
Seeing asymmetric bilayer structure with SANS
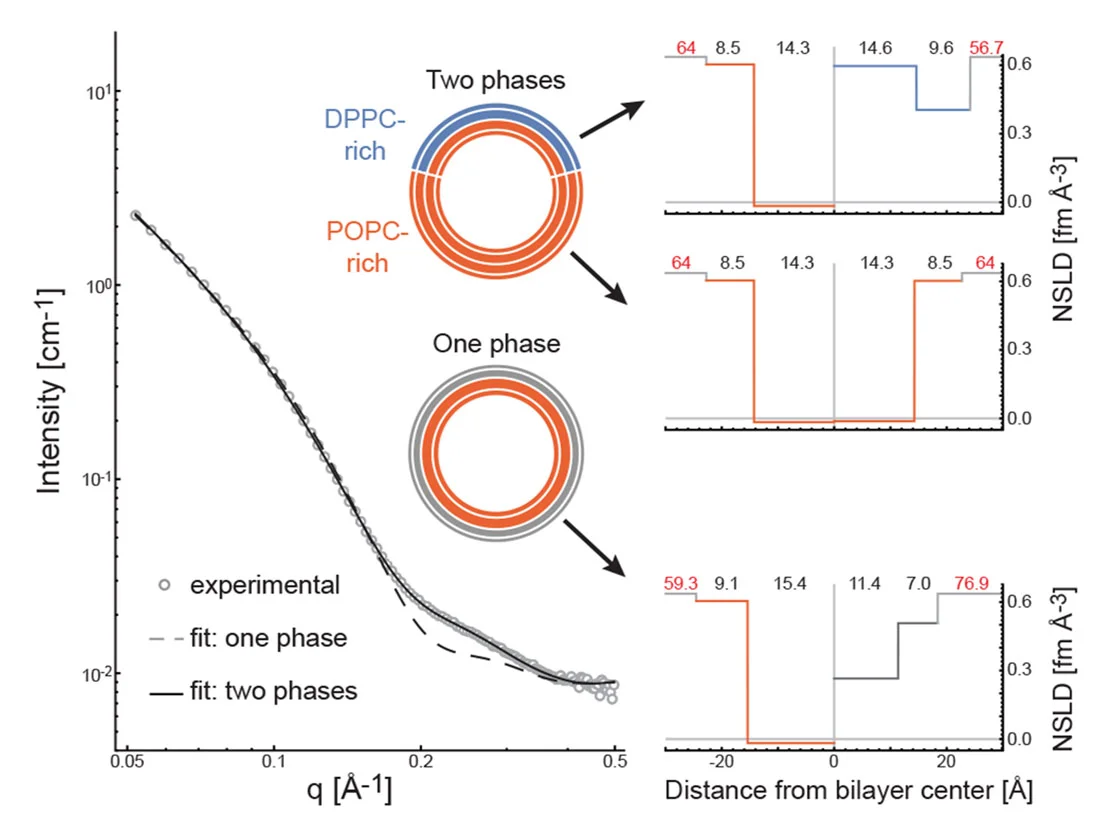
Our group specializes in the development of innovative methods for determining the structure of asymmetric membranes using small-angle neutron scattering (SANS). We use deuterated lipids to generate the contrast that allows SANS to “see” each leaflet separately. Importantly, deuterium (a stable isotope of hydrogen) is nearly identical to hydrogen in its chemistry, and replacing some of a biological molecule’s hydrogen with deuterium usually has little effect on its properties or behavior. Yet because of their different nuclei, hydrogen and deuterium interact very differently with neutrons. As a result, deuterated molecules scatter neutrons differently than their hydrogen-containing counterparts. By comparing a leaflet in an asymmetric bilayer to that of the same lipid composition but in a symmetric bilayer, we can learn how interleaflet coupling influences membrane structure.
Research articles
Eicher B, Marquardt D, Heberle FA, Letofsky-Papst I, Rechberger GN, Appavou M-S, Katsaras J, Pabst G. 2018. Intrinsic Curvature-Mediated Transbilayer Coupling in Asymmetric Lipid Vesicles. Biophysical Journal 114:146-157.
Eicher B, Heberle FA, Marquardt D, Rechberger GN, Katsaras J, Pabst G. 2017. Joint small-angle X-ray and neutron scattering data analysis of asymmetric lipid vesicles. Journal of Applied Crystallography 50:419-429.
Heberle FA, Marquardt D, Doktorova M, Geier B, Standaert RF, Heftberger P, Kollmitzer B, Nickels JD, Feigenson GW, Katsaras J, London E, Pabst G. 2016. Subnanometer Structure of an Asymmetric Model Membrane: Interleaflet Coupling Influences Domain Properties. Langmuir 32:5195-5200.
Reviews
Heberle FA, Pabst G. 2017. Complex Biomembrane Mimetics on the Sub-Nanometer Scale. Biophysical Reviews 9:353-373.
A new way of looking at lipid flip-flop
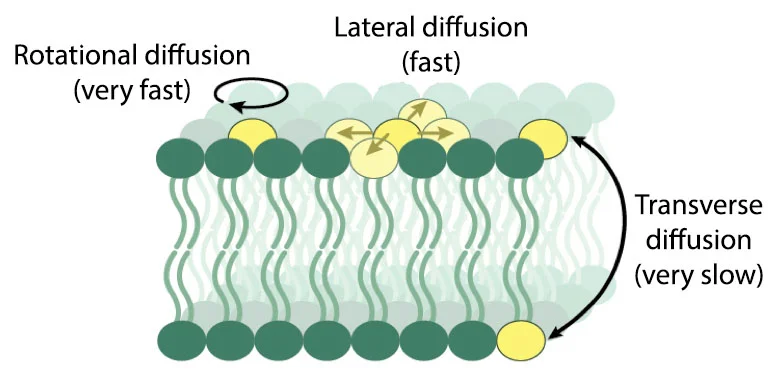
Lipids can and do move spontaneously from one bilayer leaflet to the other. The fancy word for this process is transbilayer diffusion, but these back-and-forth motions are often simply called flip-flop. Flip-flop ultimately requires moving the polar part of a lipid across the oily interior of the membrane. It’s easy to imagine that details of lipid structure—the size and charge of the headgroup, or the length and degree of unsaturation in the chains—might influence the rate at which this process occurs. Reliable values for flip-flop rates of different lipids are a necessary starting point for a detailed mechanistic understanding of plasma membrane asymmetry, yet such data are hard to come by, and the data that do exist are scattered—reported flip-flop half times range from minutes to weeks. This is in part due to the frequent use of non-native lipids tagged with a fluorophore or spin label as reporter molecules.
Asymmetric vesicles offer a unique way of measuring lipid flip-flop rates. The trick is to create a vesicle with an asymmetric distribution of a reporter lipid, and then perform some experiment that can “see” which leaflet the reporter molecule resides in. For example, proton NMR can separately resolve the inner and outer leaflet populations of lipids that have a protiated choline headgroup. If these reporters are exchanged into the outer leaflet of liposomes prepared using deuterated choline lipids, the flip-flop rate can be determined by tracking the asymmetry over time.
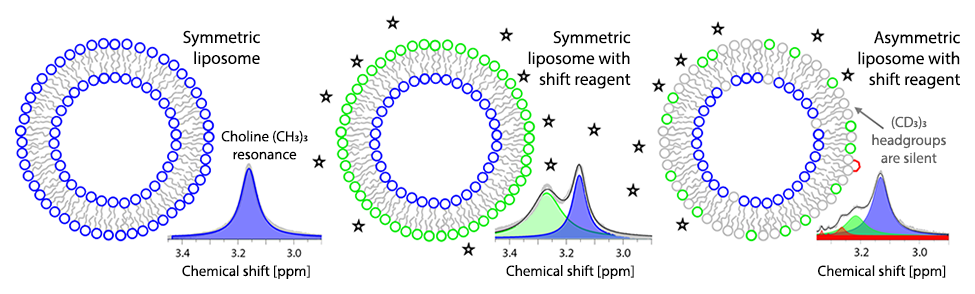
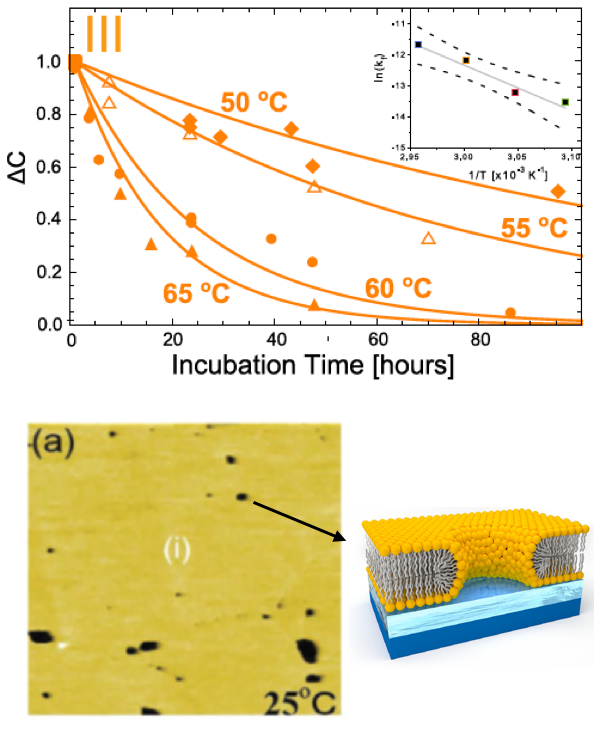
We used this method to measure the flip-flop rate of DPPC in liposomes. In the physiologically relevant fluid phase, flip-flop exhibited Arrhenius kinetics with a half time of ~4 days at 50°C—a very slow process indeed! This incredibly slow spontaneous flip-flop rate reflects the behavior of lipids in sealed vesicles that are morphologically similar to cell membranes. Remarkably, these values are slower by several orders of magnitude compared to measurements in supported lipid bilayers, where reported flip-flop half times are on the order of minutes to hours. We hypothesized that the discrepancy might be an artifact of supported bilayer sample preparation—specifically, the well-known problem of incomplete surface coverage—that can result in long-lived hydrophilic pores in the membrane. These pores provide a route for lipids to translocate from one leaflet to the other without incurring a penalty for desolvating the polar headgroup.
Research articles
Marquardt D, Heberle FA, Miti T, Eicher B, London E, Katsaras J, Pabst G. 2017. ¹H NMR Shows Slow Phospholipid Flip-Flop in Gel and Fluid Bilayers. Langmuir 33:3731-3741.
A bacterial ion channel speeds up lipid flip-flop
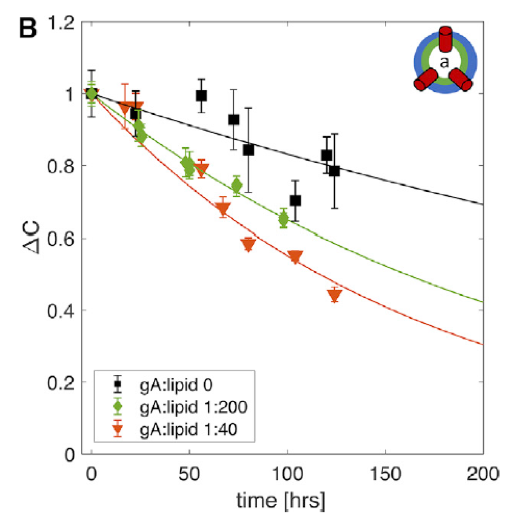
In addition to spontaneous flip-flop, proteins can accelerate lipid transbilayer movement. Some of these proteins—the so-called flippases and floppases—evolved specifically for this purpose, catalyzing the unidirectional movement of lipids from the outside in, or vice versa. Another class of transmembrane proteins called scramblases move lipids in both directions. These proteins constitute a dedicated machinery for maintaining the asymmetric lipid distribution necessary for proper cell function.
Embedded membrane proteins with completely unrelated functions are also thought to influence lipid dynamics, including their transbilayer movement. While studying the interaction of the bacterial ion channel gramicidin (gA) with asymmetric membranes, we discovered that lipid asymmetry was lost more quickly than expected. Further study with a range of techniques including small-angle X-ray scattering, differential scanning calorimetry, proton NMR, circular dichroism, and molecular dynamics simulations, revealed different mechanisms for gA-mediated lipid flip-flop depending on the protein concentration in the membrane. A computational analysis suggested that membrane deformations induced by the gA channel are in part responsible for the accelerated lipid flip-flop.
Research articles
Doktorova M, Heberle FA, Marquardt D, Rusinova R, Sanford RL, Peyear TA, Katsaras J, Feigenson GW, Weinstein H, Anderson OS. 2019. Gramicidin Increases Lipid Flip-Flop in Symmetric and Asymmetric Lipid Vesicles. Biophysical Journal 116:860-873.
Nguyen MHL, DiPasquale M, Rickeard BW, Doktorova M, Heberle FA, Scott HL, Barrera FN, Taylor G, Collier CP, Stanley CB, Katsaras J, Marquardt D. 2019. Peptide-Induced Lipid Flip-Flop in Asymmetric Liposomes Measured by Small Angle Neutron Scattering. Langmuir 35:11735-11744.
Interactive tools
Asymmetric Bilayer Phase Diagram → Compute and visualize the phase diagram of an asymmetric lipid bilayer using the Bragg–Williams mean-field model. Enter values for the Flory–Huggins interaction parameter χ and the interleaflet coupling Λ to see registered and anti-registered coexistence regions, tielines, and consolute points.